Jamie Dunning1, Anvay Patil2,7, Liliana DAlba3,6, Alexander L. Bond4,
Gerben Debruyn3, Ali Dhinojwala2, Matthew Shawkey3 and Lukas Jenni5
1Department of Life Sciences, Imperial College London, London, UK
2School of Polymer Science and Polymer Engineering, The University of Akron, Akron, OH, USA
3Department of Biology, Evolution and Optics of Nanostructure Group, University of Ghent, Gent 9000, Belgium
4Bird Group, The Natural History Museum, Tring, UK
5Swiss Ornithological Institute, Sempach, Switzerland
6Naturalis Biodiversity Center, Leiden, The Netherlands
7CertainTeed LLC, Malvern, PA, USA
JD, 0000-0001-8234-8526; AP, 0000-0003-2786-2623; LD'A, 0000-0002-2478-3455;
AD, 0000-0002-3935-7467; MS, 0000-0002-5131-8209
Publicado: 01 de Março de 2023 em https://doi.org/10.1098/rsif.2022.0920
Abstracto
Até recentemente, e quando comparada com aves diurnas que usam manchas de plumagem contrastantes e estruturas complexas de penas para transmitir informações visuais, a comunicação em espécies nocturnas e crepusculares era considerada como seguindo canais acústicos e químicos. No entanto, muitas aves que são activas em ambientes de pouca luz desenvolveram manchas de plumagem intensamente brancas dentro de plumagens que de outra forma seriam discretas. Usamos espectrofotometria, microscopia eletrónica e modelagem óptica para explicar os mecanismos que produzem as pontas das penas da cauda brancas brilhantes da galinhola eurasiática Scolopax rusticola. Sua reflectância difusa foi aproximadamente 30% maior do que qualquer pena medida anteriormente. Essa intensa reflectância é o resultado da dispersão de luz incoerente de uma nanoestrutura desordenada composta de queratina e ar dentro dos ramos da farpa. Além disso, o achatamento, o espessamento e o arranjo dessas farpas criam uma macroestrutura semelhante a uma veneziana que aumenta a área de superfície para reflexão da luz. Sugerimos que as galinholas desenvolveram essas manchas de penas brancas brilhantes para comunicação visual de longo alcance em ambientes pouco iluminados.
O uso de manchas de plumagem contrastantes ou estruturas de penas complexas para transmitir informações é amplamente difundido em aves (revisto em [1, 2]). Ao contrário das aves diurnas, os sinais visuais nas espécies noturnas e crepusculares são pouco estudados, e a comunicação era, até recentemente, considerada como seguindo canais químicos e acústicos [3 - 5]. No entanto, em ambientes com pouca luz, surgiram características de plumagem que maximizam a reflectância da luz disponível [6, 7]. Embora a maioria das aves noturnas e crepusculares tenham plumagens discretas ou enigmáticas, os sinais visuais são tipicamente de um branco intenso; por exemplo, as manchas brancas na plumagem de alguns noitibós Caprimulgidae [8], corujas Strigidae [9 - 11], maçaricos Burhinidae [12] e narcejas Scolopacidae [13].
A função e o mecanismo pelo qual essas manchas brancas optimizam a reflectância da luz não são bem compreendidos (veja [14,15]), mas provavelmente comunicam intenção, por exemplo, acasalamento ou comportamentos territoriais ou qualidade do sinal ([13]; ver também [16]). No entanto, as aves noturnas e crepusculares normalmente também requerem crípse durante o dia [17,18] e, portanto, ocultam seus sinais visuais. Manchas brancas nas asas de alguns noitibós são, por exemplo, expostas apenas em voo [8]; Ou, nas galinholas Scolopax spp, manchas de penas brancas na parte inferior da cauda só são expostas quando a cauda é levantada, ou quando se envolve em vôos de demonstração ([19]; figura 2).
Borodulina & Formosow [19] descreveram pela primeira vez modificações nos ramos (que irradiam da raque central da pena) que compreendem as pontas brancas na parte inferior das penas da cauda (doravante retrizes) da galinhola eurasiática Scolopax rusticola (doravante galinhola; figura 2 a ), mas não mediu a refletância e caracterizou o seu mecanismo. Estudos anteriores demonstraram como as microestruturas se correlacionam com a intensidade da plumagem branca, por exemplo, na plumagem do corpo de inverno do tetraz-das-rochas Lagopus muta [20], as cores semelhantes a opalas em alguns pássaros piprídeos Pipridae [14] e entre muitos pássaros de plumagem branca de famílias diferentes [15]. Da mesma forma, 'super-branco', derivado de microestruturas na carapaça de um besouro [21, 22] foram bem relatados. As manchas brancas em aves noturnas e crepusculares, que são potencialmente optimizadas para sinalização em condições de pouca luz, raramente foram abordadas e requerem uma análise mais detalhada.
Aqui descrevemos os mecanismos pelos quais as rectrizes brancas da galinhola produzem um sinal branco intenso em condições de pouca luz, usando espectrofotometria difusa e resolução de ângulo, microscopia eletrónica e modelagem óptica por meio de abordagens de domínio do tempo de diferenças finitas (FDTD).
Para caracterizar a microestrutura e a nanoestrutura responsáveis ??pela produção do sinal branco brilhante, utilizamos microscopia eletrónica de varredura e transmissão (SEM e TEM, respectivamente). Para SEM, montamos ramos individuais brancos e castanhos (obtidos da mesma pena) separadamente, em tocos com fita de carbono. Também orientamos pequenos fragmentos de ramos de forma que permitisse a sua observação em corte transversal. Revestimos as amostras com ouro/paládio por 2 minutos e as visualizamos em um SEM (FlexSEM 1000; Hitachi) a uma tensão de aceleração de 10 kV e 6 mm de distância de trabalho.
Para o TEM, primeiro incorporamos rami individuais seguindo um protocolo padrão [23]. Resumidamente, enxaguamos e desidratamos os rami com etanol três vezes, e depois infiltramo-los com concentrações crescentes (15%, 50%, 70% e 100%) de resina epóxi (EMbed-812; Electron Microscopy Sciences, PA, EUA) seguidos de 16 horas de polimerização em resina epóxi a 60°C em estufa de laboratório.
Aparamos os blocos contendo os ramos e cortamos seções transversais de 100 nm de espessura usando um ultramicrótomo Leica UC-6 (Leica Microsystems, Alemanha). Recolhemos as secções usando carbono com fenda oval e grades de cobre revestidas com formvar em duplicata e coradas com Uranyless/citrato de chumbo. Observamos as secções em um microscópio eletrônico de transmissão JEOL JEM 1010 (Jeol Ltd, Tóquio, Japão) operando a 120 kV.
Usamos micro e (macro) espectrofotometria para medir espectros de refletância de três rectrizes separadas. Usamos as mesmas penas, mas de aves diferentes para todas as análises. Medimos a refletância da superfície reversa (lado inferior) de um ramo branco usando um microespectrofotômetro (CRAIC AX10: sensibilidade 320800 nm) e um espectrofotômetro que mediu uma região em vários ramos (aproximadamente 2 mm de tamanho do ponto). Medimos a refletância difusa (toda a luz reflectida) e especular (luz refletida num ângulo específico) entre 300 e 700 nm em incrementos de 1 nm usando um espectrômetro AvaSpec-2048 e configuração de fonte de luz dupla (AvaLight-DH-S deutério-fonte de luz halógena e fonte de luz AvaLight-HAL-S-MINI). Medimos a refletância difusa (que assume que a refletância da luz é influenciada por estruturas internas, bem como aquelas na superfície de um objeto) usando uma sonda bifurcada e uma esfera integradora com uma armadilha de brilho preto para excluir a refletância especular (luz refletida da superfície de um objeto) ( AvaSphere-50-REFL). Em seguida, medimos a refletância especular em três ângulos diferentes (75°, 60°, 45°) usando uma sonda bifurcada e um porta-bloco (AFH-15, Avantes). Colocamos cada pena em papel preto minimizando a refletância do fundo. Todas as medições são expressas em relação a um padrão de refletância branca de 99% (WS-2, Avantes) e padrão preto Avantes de 2% (BS-2, Avantes). Processamos dados no pacote R pavo em R 4.1.2 [ s superfície) usando uma sonda bifurcada e uma esfera integradora com uma armadilha de brilho preto para excluir a refletância especular (luz refletida da superfície de um objeto) (AvaSphere-50-REFL). Em seguida, medimos a refletância especular em três ângulos diferentes (75°, 60°, 45°) usando uma sonda bifurcada e um porta-bloco (AFH-15, Avantes). Colocamos cada pena em papel preto minimizando a refletância do fundo. Todas as medições são expressas em relação a um padrão de refletância branca de 99% (WS-2, Avantes) e padrão preto Avantes de 2% (BS-2, Avantes). Processamos dados no pacote R pavo em R 4.1.2 [ s) usando uma sonda bifurcada e uma esfera integradora com uma armadilha de brilho preto para excluir a refletância especular (luz refletida da superfície de um objeto) (AvaSphere-50-REFL). Em seguida, medimos a refletância especular em três ângulos diferentes (75°, 60°, 45°) usando uma sonda bifurcada e um porta-bloco (AFH-15, Avantes). Colocamos cada pena em papel preto minimizando a refletância do fundo. Todas as medições são expressas em relação a um padrão de refletância branca de 99% (WS-2, Avantes) e padrão preto Avantes de 2% (BS-2, Avantes). Processamos dados no pacote R pavo em R 4.1.2 [ Colocamos cada pena em papel preto minimizando a refletância do fundo. Todas as medições são expressas em relação a um padrão de refletância branca de 99% (WS-2, Avantes) e padrão preto Avantes de 2% (BS-2, Avantes). Processamos dados no pacote R pavo em R 4.1.2 [ Colocamos cada pena em papel preto minimizando a refletância do fundo. Todas as medições são expressas em relação a um padrão de refletância branca de 99% (WS-2, Avantes) e padrão preto Avantes de 2% (BS-2, Avantes). Processamos dados no pacote R pavo em R 4.1.2 [24, 25] e comparamos com medições publicadas anteriormente de 61 outras aves usando métodos espectrofotométricos idênticos [15].
Para explorar a direccionalidade da refletância em função da variação do ângulo de rami, modelamos como os fótons interagem com as estruturas dentro de uma farpa individual. Executamos uma série de simulações de domínio do tempo de diferenças finitas (FDTD) usando um solucionador Ansys Lumerical 2021 R1 de nível comercial (Ansys, Inc.). O método FDTD fornece uma solução geral para qualquer problema de dispersão de luz em geometrias arbitrárias complexas (neste caso, uma estrutura de 'célula unitária' renderizada por desenho assistido por computador (CAD) de um ramo individual) resolvendo numericamente as equações de rotação de Maxwell numa estrutura discreta grade espaço-temporal [26]. A simulação estima toda a luz espalhada em um ângulo de incidência específico (daqui em diante, AOI). Esses resultados simulados podem ser comparados com os dados de espectrofotometria difusa.
Nossos modelos CAD tridimensionais simulados foram baseados em observações microscópicas empíricas das farpas da galinhola (ver material eletrónico suplementar, S1:AD). Primeiro, renderizamos uma geometria CAD tridimensional para uma célula unitária oca de controle, sem nanoestruturas fotônicas internas (neste caso, a geometria sem as estruturas observadas na figura 1 h ) e uma célula unitária de controle sólida.
Figura 1 (ver na capa da notícia)
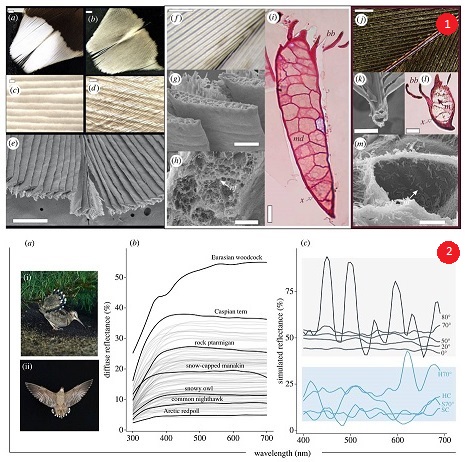
Figura 1. ( a )( e ): Morfologia das pontas brancas das rectrizes da Scolopax rusticola. ( a ) Superfície reversa branca. ( b ) Superfície anverso castanha. ( c ) Rami branco em alinhamento veneziano; células individuais são aparentes. ( d ) Vista anversa mostrando as bárbulas escuras entrelaçadas cobrindo os ramos brancos. ( e ) Micrografia SEM da ponta da rectriz branca cortada transversalmente, mostrando a superfície rasa em forma de V dos ramos; ( f )( m ): Comparação da microestrutura das partes brancas e castanhas das rectrizes. ( f ) Imagem óptica de ramos brancos com aumento de 30x. (g ) Ramos engrossados ??e achatados vistos da superfície reversa. ( h ) O interior de um ramo branco mostra células com redes de fibras de queratina ( nf ) e bolsas de ar. ( i ) um ramo branco mostrando células medulares ocas ( md ) e um córtex fino ( x ); as bárbulas ( bb ) estão presentes no lado anverso. ( j ) Imagem óptica da região castanha contígua com aumento de 30x. ( k ) Ramos castanhos em corte transversal. ( l ) Melanossomas ( m ) presentes ao longo dos ramos e bárbulas. ( m ) Célula medular do ramo castanho mostrando melanossomas ( m) e a ausência de matrizes de queratina. Barras de escala: ( a ) e ( b ) 1 mm; (c), ( d ), ( g ) e ( k ) 50 µm; ( e ) 500 ?m; ( h ) 10 ?m; ( i ) e ( l ) 100 ?m; ( m ) 5 ?m; ( f ) e ( j ) 1 mm.
Usamos microscopia SEM para definir as dimensões do CAD e definimos cada célula unitária por uma espessura do córtex de queratina de 7 µm com um interior oco, 20 µm de altura (direção Z) e 8 µm de largura (direcção X). Em seguida, usamos a microscopia SEM para renderizar uma célula unitária simples análoga (doravante célula unitária) com uma nanoestrutura interna equivalente aos ramos da galinhola, ou seja, de bolsas de ar e uma matriz de suporte de nanofibras (figura 1). Fizemos isso usando uma distribuição aleatória uniforme de partículas esféricas não sobrepostas dentro da matriz de queratina, que variaram aleatoriamente em diâmetro entre 0,45 e 3,45 µm. As constantes ópticas (índices de refração complexos) para a queratina foram adaptadas da literatura anterior ([27]; material eletrônico suplementar, tabela S1).
Realizamos simulações usando uma fonte de onda plana de banda larga (400700 nm), propagada ao longo da direção ?Z. Primeiro, em um AOI normal (0°) e depois a 70° da superfície normal, para nossas células unitárias de controle, ocas e sólidas, renderizadas em CAD. Em seguida, executamos simulações usando nossa célula unitária de galinhola simulada a 0° e a 20°, 50°, 70° e 80° AOI da superfície normal. As condições de contorno na direção lateral (X e Y) foram definidas como periódicas. Monitorizamos os dados de refletância usando um monitor de energia de transformação discreta de Fourier (DFT) colocado atrás do plano de injeção da fonte. O tempo de simulação (em fs) e a condição de contorno ao longo da direção de propagação da luz (Z; limites de camada perfeitamente correspondentes (PML)) foram escolhidos de forma que o campo eléctrico decaísse antes do final da simulação (critérios de desligamento automático). Toda a luz incidente foi refletida, transmitida ou absorvida.
2.4. Prevalência em táxons relacionados
Finalmente, examinamos espécimes de oito espécies de galinhola (incluindo S. rusticola ) e vinte e três espécies de Scolopacidae não Scolopax estreitamente relacionadas em coleções de museus para pontas de penas de cauda branca, combinando com as descritas aqui (como figura 2 a ; ver Agradecimentos para uma lista de instituições).
Figura 2 (ver na capa da notícia)
Figura 2. ( a ) Galinhola Eurasiana Scolopax rusticola mostrando o contexto ecológico quando as pontas brancas são expostas, seja do solo (provavelmente uma fêmea atraindo um macho sobrevoando) ( a (i)) ou em voo (macho em voo de exibição) ( a ( ii)); fotos de Serge Santiago e Jean-Lou Zimmermann. ( b ) Espectros de refletância difusa medidos a partir da superfície reversa das pontas brancas das rectrizes da galinhola da Eurásia, com pico de aproximadamente 55%, 31% mais brilhante do que a próxima pena mais brilhante, Caspian tern Hydroprogne caspia, e comparado com 61 plumagens brancas de Igic et al. [15], as espécies citadas no texto são destacadas; ( c) simulações de domínio do tempo de diferenças finitas (FDTD) mostrando refletância simulada em cinco ângulos de incidência de uma célula unitária renderizada por CAD representando a ponta de uma pena de cauda de galinhola (AOI; destacada em cinza, 0° (superfície normal), 20°, 50°, 70° e 80°) e duas células unitárias de controle, medidas em AOI (destacadas em azul; uma célula unitária oca a 70° (H70°), um controle oco na superfície normal (HC), controle sólido a 70° (S70°) e controle sólido na superfície normal (SC)). Esses dados sugerem que os bolsões de ar presentes na matriz de queratina são essenciais para aumentar a refletância nos comprimentos de onda visíveis nas penas da cauda da galinhola.
3.1. Estrutura das pontas rectrix brancas
As pontas das rectrizes são brancas no reverso (figuras 1 a e 2 a ), mas castanhas acinzentadas na superfície do anverso (face superior) (figura 1 b ). Os ramos são engrossados ??e achatados na mancha branca e sobrepostos uns aos outros, superficialmente como venezianas (figura 1 c,e ). O ângulo desses ramos em relação à superfície da pena varia (conforme sugerido em [19]), estimamos de aproximadamente 70° para os ramos proximais a aproximadamente 76° para os ramos distais (figura 1 e). As bárbulas proximais e distais castanhas originam-se da superfície superior dos ramos, portanto, são visíveis apenas na superfície anverso e cobrem os ramos brancos espessados ????por cima, fornecendo a coloração castanha acinzentada da superfície anverso (figura 1 b, d ) . Eles entrelaçam-se para formar uma palheta coerente. Os dois lados de uma ponta branca, separados pela raque, são côncavos e as farpas dispostas em ângulos opostos (figura 1 j), refletindo a luz em diferentes direções e aparentes ao virar uma pena em pouca luz. Em contraste, as partes castanhas das rectrizes são estruturalmente típicas de penas de palhetas com farpas finas que são espaçadas pelas bárbulas castanhas (figura 1 j , k). Os ramos brancos espessos nas pontas das penas eram aproximadamente 2,5 vezes mais espessos e pareciam internamente mais complexos do que os ramos castanhos (figura 1 fh e jm , respectivamente). A medula dos ramos brancos continha numerosas e complexas células fotônicas com finas redes de nanofibras e bolsões de ar dispersos (figura 1 gi), totalmente sem melanossomas. Essas matrizes de ar e queratina pareciam desorganizadas. Em contraste, os ramos das regiões de penas castanhas eram menos espessos, mais redondos, tinham menos células medulares e não continham uma matriz de ar e queratina, mas eram abundantes em melanossomas tanto dentro da medula da farpa quanto no córtex (figura 1 k- m) .
3.2. Refletância
A espectrofotometria revelou intensa reflectância difusa através dos ramos na parte inferior branca das retrizes, com pico de 55% (628 nm) (figuras 1 f e 2 a ). Da mesma forma, ramos individuais tiveram refletância especular ainda maior, com pico superior a 100% contra um padrão difuso (material eletrónico suplementar, figura S2). As manchas brancas nas retrizes da galinhola são, portanto, excepcionalmente brilhantes e, até onde sabemos, representam o branco mais brilhante medido a partir da plumagem de um pássaro, 31% mais brilhante do que o próximo mais reflexivo, Caspian tern Hydroprogne caspia , que atinge o pico em 38 % (459 nm) e 91% mais brilhante do que a pena branca menos reflexiva medida, Arctic redpoll Acanthis hornemanni, que atinge o pico em 4,9% (638 nm) ([15]; figura 2 b ). A reflexão especular foi maior quando medida a 75° em relação à superfície normal, diminuindo em ângulos mais agudos (material eletrónico suplementar, figura S3).
3.3. Simulações de refletância no domínio do tempo com diferenças finitas
Descobrimos que a nanoestrutura desordenada formada por queratina e fases de ar nos ramos da galinhola foram essenciais para gerar intensa refletância branca. Para incidência normal, a refletância geral (integrada em todos os ângulos) para a nanoestrutura de célula unitária de rami imitada por galinhola aumentou aproximadamente 65% em relação à nanoestrutura de célula unitária oca de controle. Além disso, as simulações também destacam alguma direcionalidade para a intensidade do patch. A refletância modelada a 80° resultou em maior oscilação nos espectros, derivada de interferência construtiva e destrutiva. Caso contrário, a refletância aumentou de um pico de aproximadamente 45% na incidência normal para um pico de aproximadamente 57% a 70°, que representa o ângulo real dos ramos dentro da mancha branca (figura 2 c). A refletância modelada em 75° (não mostrada na figura 2 c , mas veja [28]; material eletrónico suplementar, figura S4) e 80° AOI mostraram ruído crescente, que sugerimos ser devido a efeitos de interferência (destacados por desvios padrão crescentes em torno da média refletância à medida que AOI aumenta; material eletrônico suplementar, figura S4). A refletância a 70° é basicamente a mesma que a refletância difusa real (figura 2 c), embora o FDTD simule refletância difusa mais especular. Portanto, sugerimos que os ramos sejam dispostos no ângulo que melhor otimiza a refletância. Além disso, nossas células de unidade de controle simuladas demonstram que os bolsões de ar na matriz de queratina são essenciais para aumentar a refletância geral nos comprimentos de onda visíveis.
3.4. Prevalência em táxons relacionados
Registramos manchas brancas equivalentes, definidas pela presença de ramos brancos espessos e achatados nas pontas das penas da cauda em todas as oito espécies de galinhola, mas não em seus parentes mais próximos (23 espécies não Scolopax Scolopacidae, ver material eletrónico suplementar, tabela S1).
Nossos resultados sugerem que as pontas brancas nas rectrizes da galinhola representam a refletância mais brilhante já medida e, em virtude, a mancha de plumagem branca mais branca atualmente conhecida entre as aves. Embora intensa refletância em plumagens brancas tenha sido relatada anteriormente [20, 29, 30], elas não tiveram comparação padronizada com outras espécies de plumagem branca. Assim, apresentamos nossos resultados juntamente com as plumagens descritas anteriormente (ver [15] para uma lista completa), usando métodos espectrofotométricos padronizados (figura 2 b). Essa reflexão é produzida pela disposição de ramos grossos e achatados com ampla distribuição de bolsas de ar, que juntas maximizam a intensidade da reflexão da luz. Usamos simulações FDTD para demonstrar que (i) a estrutura interna dos ramos nas pontas brancas é integral para dispersão de luz e subsequente intensidade de reflexão, mas também (ii) que o ângulo das farpas ampliadas em relação umas às outras optimiza a refletância em a macroescala.
As estruturas que descrevemos diferem daquelas de plumagens brancas reflexivas menos intensas de duas maneiras: primeiro, os ramos são engrossados ??e achatados ([19]; este estudo), aumentando a área de superfície disponível para reflexão e impedindo a passagem de luz entre os ramos e as bárbulas . Em segundo lugar, os ramos espessos permitem uma complexidade de células fotônicas, com uma rede de nanofibras de queratina e bolsas de ar espalhadas, criando inúmeras interfaces para favorecer eventos de espalhamento (como a refletância 'superbranca' descrita em um besouro branco; [21, 22]).
Igic et al. [15] sugeriram que a refletância mais intensa da plumagem branca estava associada a ramos densamente compactados, mais redondos e menos ocos, mas também bárbulas mais espessas e longas. Consequentemente, as espécies maiores eram mais brilhantes em virtude da espessura e complexidade dos ramos. No entanto, os ramos da galinhola são engrossados ??e achatados, superficialmente como os ramos da coroa branca do tangará-azul Lepidothrix isidorei [14]; neste caso, a nanoestrutura interna está sem os ramos espessados ??que aumentam a área de superfície de reflexão. Apesar de algumas semelhanças, a reflectância difusa da coroa do tangará atinge um pico de aproximadamente 17% [14], aproximadamente 105% menos brilhante que a galinhola. No entanto, a refletância especular da coroa do tangará é maior do que a da galinhola, devido a uma nanoestrutura que aumenta a refletância especular (ver também [31, 32]). O arranjo veneziano dos ramos espessos e a subsequente reflexão direcional é superficialmente como o arranjo das bárbulas de alguns beija-flores Trochilidae. Neste caso, o ângulo das bárbulas em relação ao eixo do ramo, e o ângulo entre as bárbulas proximal e distal dos ramos determinam a direcionalidade da refletância, associada à irradiância [ 33 ], semelhante ao mecanismo que descrevemos na ponta da pena da cauda da galinhola.
Manchas brancas estão presentes em todas as oito espécies de galinhola, mas não em seus parentes mais próximos (23 espécies não Scolopax Scolopacidae, ver material eletrónico suplementar, tabela S1). Não realizamos medições de espectrofotometria nem trabalho de microscopia em nenhum táxon de galinhola, além de S. rusticola , e assim não pudemos comparar seus respectivos espectros de reflectância.
Sugerimos que as manchas brancas presentes nas penas da cauda das galinholas estão ligadas à sinalização de algum comportamento em ambientes pouco iluminados [12, 34]. Como essas manchas são visíveis apenas por baixo, qualquer significado funcional depende de levantar e abanar a cauda, ??por exemplo, durante exibições de corte [35 - 38], distração do predador ou comunicação não reprodutiva [39, 40]. A ligação entre a intensidade do patch, o comportamento e o ambiente de luz relativa é pouco estudada e beneficiaria de mais pesquisas.
Sugerimos que as galinholas desenvolveram manchas de penas brancas brilhantes, as mais brilhantes descritas nas aves, por meio de modificações estruturais elaboradas nas escalas macro, micro e nano para comunicação em ambientes pouco iluminados.
Ética
Nenhum animal foi ferido ou sacrificado para os propósitos deste estudo. As amostras de penas da cauda da galinhola, utilizadas nos experimentos aqui apresentados, foram de muda de aves (coletadas e armazenadas por Niklaus Zbinden, Suíça, 1995), sem a necessidade de licenciamento específico. Todos os procedimentos experimentais foram realizados de acordo com as políticas de ética das instituições de pesquisa afiliadas.
Acessibilidade de dados
Os dados originais coletados como parte deste estudo foram disponibilizados no Repositório Digital Dryad [28].
Os dados são fornecidos em material eletrônico suplementar [41].
Contribuições dos autores
JD: conceituação, curadoria de dados, análise formal, investigação, metodologia, administração do projeto, recursos, supervisão, validação, visualização, redaçãorascunho original, redaçãorevisão e edição; AP: conceituação, análise formal, metodologia, visualização, redaçãorascunho original, redaçãorevisão e edição; LD: conceituação, análise formal, investigação, metodologia, recursos, validação, visualização, redaçãorascunho original, redaçãorevisão e edição; ALB: curadoria de dados, investigação, metodologia, redaçãorascunho original, redaçãorevisão e edição; GD: análise formal, investigação, validação, redação revisão e edição; AD: conceituação, análise formal, obtenção de financiamento, investigação, recursos, supervisão, redação revisão e edição; MS: conceituação, curadoria de dados, análise formal, obtenção de financiamento, investigação, metodologia, administração do projeto, supervisão, validação, redaçãorascunho original, redaçãorevisão e edição; LJ: conceituação, curadoria de dados, análise formal, investigação, metodologia, administração do projeto, supervisão, validação, redaçãorascunho original, redaçãorevisão e edição.
Todos os autores deram a aprovação final para publicação e concordaram em ser responsabilizados pelo trabalho realizado.
Declaração de conflito de interesses
Declaramos que não temos interesses concorrentes.
Financiamento
JD foi apoiado pelo QMEE CDT, financiado pelo NERC (bolsa nº NE/P012345/1); AD, AP e MS reconhecem o apoio financeiro do Escritório de Pesquisa Científica da Força Aérea dos EUA (AFOSR) sob a concessão da Iniciativa de Pesquisa Universitária Multidisciplinar (MURI) (concessão nº FA 9550-18-1-0142) e concessão suplementar FA-9550-18 -1-0477. A MS também reconhece a concessão RGP0047 do Programa de Pesquisa Human Frontiers e a concessão G007117N dos Fundos de Pesquisa Flamenga (FWO).
Reconhecimentos
Gostaríamos de agradecer a Mary Hennen (Field Museum, Chicago), Michael A. Pearson (South Notts Ringing Group), Matt Rayner (Auckland Museum), Julia Schroeder (Imperial College London), Paul Sweet (American Museum of Natural History), Arseny Tsvey (Academia Russa de Ciências), Niklaus Zbinden (Instituto Ornitológico Suíço) e aqueles que nos enviaram penas de galinhola, para conversa, espécimes ou traduções. Agradecemos também a Serge Santiago e Jean-Lou Zimmermann pelo uso das fotos da figura 2 a .
Notas de rodapé
O material suplementar eletrônico está disponível online em https://doi.org/10.6084/m9.figshare.c.6430381 .
© 2023 Os Autores.
Publicado pela Royal Society sob os termos da Creative Commons Attribution License http://creativecommons.org/licenses/by/4.0/ , que permite o uso irrestrito, desde que o autor original e a fonte sejam creditados.
1. Jenni L, Winkler R. 2020 The biology of moult
in birds. London, UK: Helm.
2. Terril RS, Shultz AJ. 2022 Feather function
and the evolution of birds. Biol. Rev. (doi:10.1111/
brv.12918)
3. Healy S, Guilford T. 1990 Olfactory-bulb size and
nocturnality in birds. Evolution 44, 339346.
(doi:10.2307/2409412)
4. Bonadonna F, Bretagnolle V. 2002 Smelling home: a
good solution for burrow-finding in nocturnal
petrels? J. Exp. Biol. 205, 25192523. (doi:10.1242/
jeb.205.16.2519)
5. Grieves LA, Gilles M, Cuthill IC, Székely T,
MacDougall-Shackleton EA, Caspers BA. 2022
Olfactory camouflage and communication in
birds. Biol. Rev. 97, 11931209. (doi:10.1111/
brv.12837)
6. Endler JA. 1993 The color of light in forests and its
implications. Ecol. Monogr. 63, 127. (doi:10.2307/
2937121)
7. Penteriani V, Del Mar Delgado M. 2017 Living in the
dark does not mean a blind life: bird and mammal
visual communication in dim light. Proc. R. Soc. B
372, 20160064. (doi:10.1098/rstb.2016.0064)
8. Aragonés J, Arias De Reyna L, Recuerda P. 1999
Visual communication and sexual selection in a
nocturnal bird species, Caprimulgus ruficollis, a
balance between crypsis and conspicuousness.
Wilson Bull. 111, 340345.
9. Penteriani V, Del Mar Delgado M, Alonso-Alvarez C,
Sergio F. 2007 The importance of visual cues for
nocturnal species: eagle owls signal by badge
brightness. Behav. Ecol. 18, 143147. (doi:10.1093/
beheco/arl060)
10. Bortolotti GR, Stoffel MJ, Galván I. 2011 Wintering
snowy owls Bubo scandiacus integrate plumage
colour, behaviour and their environment to
maximize efficacy of visual displays. Ibis
153, 134142. (doi:10.1111/j.1474-919X.2010.
01067.x)
11. Bettega C, Campioni L, Del Mar Delgado M,
Lourenço R, Penteriani V. 2013 Brightness features
of visual signalling traits in young and adult
Eurasian eagle-owls. J. Raptor Res. 47, 197207.
(doi:10.3356/JRR-12-00002.1)
12. Cramp S, Simmons KEL. 1983 Handbook of the birds
of Europe, the Middle East and North Africa. Vol 3:
The birds of the Western Palearctic. Oxford, UK:
Oxford University Press.
13. Höglund J, Eriksson M, Lindell LE. 1990
Females of the lek-breeding great snipe,
Gallinago media, prefer males with white tails.
Anim. Behav. 40, 2332. (doi:10.1016/S0003-
3472(05)80662-1)
14. Igic B, DAlba L, Shawkey MD. 2016 Manakins can
produce iridescent and bright feather colours
without melanosomes. J. Exp. Biol. 219,
18511859. (doi:10.1242/jeb.137182)
15. Igic B, DAlba L, Shawkey MD. 2018 Fifty shades of
white: how white feather brightness differs among
species. Sci. Nat. 105, 34. (doi:10.1007/s00114-
018-1543-3)
16. Sæther SA, Fiske P, Kålås JA, Gjul JM. 2000 Females
of the lekking great snipe do not prefer males with
whiter tails. Anim. Behav. 59, 273280. (doi:10.
1006/anbe.1999.1301)
17. Troscianko J, Wilson-Aggarwal JK, Stevens M,
Spottiswoode CN. 2016 Camouflage predicts survival
in ground-nesting birds. Sci. Rep. 6, 19966. (doi:10.
1038/srep19966)
18. Stevens M, Troscianko J, Wilson-Aggarwal JK,
Spottiswoode CN. 2017 Improvement of individual
camouflage through background choice in groundnesting
birds. Nat. Ecol. Evol. 1, 13251333. (doi:10.
1038/s41559-017-0256-x)
19. Borodulina TL, Formosow AN. 1967 About signal
spots of feathering of birds and peculiarity of
woodcock rectrices (in Russian). Bjull. Mosk. obstsch.
ispyt. prirody, otd. biol. 72, 2731. Also published
in Russkii Ornitologicheskii Zhurnal 24. 2015.
Ekspress Vypusk 1230: 46224626.
20. Dyck J. 1979 Winter plumage of the rock
ptarmigan: structure of the air-filled barbules and
function of the white colour. Dansk. Orn. Foren.
Tidsskr. 73, 4158.
21. Vukusic P, Hallam B, Noyes J. 2007 Brilliant
whiteness in ultrathin beetle scales. Science 315,
348. (doi:10.1126/science.1134666)
22. Burresi M, Cortese L, Pattelli L, Kolle M, Vukusic P,
Wiersma DS, Steiner U, Vignolini S. 2014 Brightwhite
beetle scales optimise multiple scattering of
light. Sci. Rep. 4, 6075. (doi:10.1038/srep06075)
23. DAlba L, Meadows M, Maia R, Jong-Souk Y,
Manceau M, Shawkey M. 2021 Morphogenesis of
iridescent feathers in Annas hummingbird Calypte
anna. Integr. Comp. Biol. 61, 15021510. (doi:10.
1093/icb/icab123)
24. Maia R, Gruson H, Endler JA, White TE. 2019 pavo
2: new tools for the spectral and spatial analysis of
colour in R. Methods Ecol. Evol. 10, 10971107.
(doi:10.1111/2041-210x.13174)
25. R Core Team. 2022 R: A language and environment
for statistical computing. Vienna, Austria: R
Foundation for Statistical Computing. See https://
www.R-project.org/.26. Taflove A, Hagness SC. 2005 Computational
electrodynamics: the finite-difference time-domain
method, 3rd edn. Norwood, MA: Artech House, Inc.
27. Stavenga DG, Leertouwer HL, Osorio DC, Wilts BD.
2015 High refractive index of melanin in shiny
occipital feathers of a bird of paradise. Light Sci.
Appl. 4, e243. (doi:10.1038/lsa.2015.16)
28. Dunning J, Patil A, DAlba L, Bond AL, Debruyn G,
Dhinojwala A, Shawkey M, Jenni L. 2023 Data from:
How woodcocks produce the most brilliant white
plumage patches among the birds. Dryad Digital
Repository. (doi:10.5061/dryad.31zcrjdqs)
29. Tickell WLN. 2003 White plumage. Waterbirds: Int.
J. Waterbird Biol. 26, 112.
30. Stoddard MC, Prum RO. 2011 How colorful are birds?
Evolution of the avian plumage color gamut. Behav.
Ecol. 22, 10421052. (doi:10.1093/beheco/arr088)
31. Shawkey MD, Maia R, DAlba L. 2011 Proximate
bases of silver color in anhinga (Anhinga anhinga)
feathers. J. Morphol. 272, 13991407. (doi:10.1002/
jmor.10993)
32. McCoy DE, Shultz AJ, Vidoudez C, van der Heide E,
Dall JE, Trauger SA, Haig D. 2021 Microstructures
amplify carotenoid plumage signals in tanagers. Sci.
Rep. 11, 120. (doi:10.1038/s41598-020-79139-8)
33. Giraldo M, Sosa J, Stavenga D. 2021 Feather
iridescence of Coeligena hummingbird species
varies due to differently organized rami and
barbules. Biol. Lett. 17, 20210190. (doi:10.1098/
rsbl.2021.0190)
34. Glutz Von Blotzheim UN, Bauer KM, Bezzel E. 1977
Handbuch der vögel mitteleuropas, vol 7. Leipzig,
Germany: Akademische Verlagsgesellschaft.
35. Hagen Y. 1950 How a woodcock (Scolopax rusticola)
sitting on the ground attracts the attention of
partners in mating flight. Vår Fågelvärld 9,
195199.
36. Hirons G. 1980 The significance of roding by
woodcock Scolopax rusticola: an alternative
explanation based on observations of marked birds.
Ibis 122, 350354. (doi:10.1111/j.1474-919X.1980.
tb00888.x)
37. Ferrand Y, Gossmann F. 2009 La bécasse des bois.
Saint-Lucien, France: Effet de lisière éditeur.
38. Lastukhin AA, Isakov AM. 2016 Reflection of light
from white spots on the tail of a woodcock Scolopax
rusticola in flight (in Russian). Russkii
Ornitologicheskii Zhurnal 25, Ekspress Vypusk. 1312,
26422643.
39. Ingram C. 1974 Possible functions of the tail spots in
the woodcock. Brit. Birds 67, 475476.
40. Fetisov CA. 2017 On the functional significance of
bright white spots on the tail of the woodcock
Scolopax rusticola (in Russian). Russkii Ornitologicheskii
Zhurnal 26, Ekspress Vypusk. 1466, 27272733.
41. Dunning J, Patil A, DAlba L, Bond AL, Debruyn G,
Dhinojwala A, Shawkey M, Jenni L. 2023 How
woodcocks produce the most brilliant white
plumage patches among the birds. Figshare.
(doi:10.6084/m9.figshare.c.6430381)
Fonte: The Royal Society Publishing
Tradução: André Verde